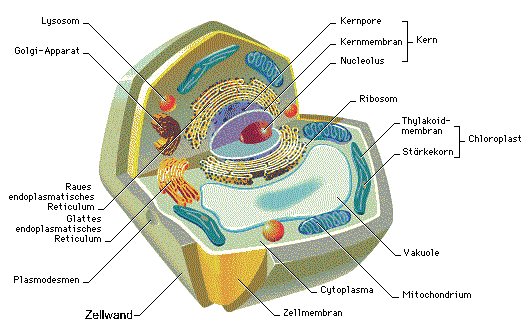
Heute möchte ich mich mit der Transkription und der Translation beschäftigen. Betrachten wir wieder die DNA als Bauplan, so ist die Transkription die Übersetzung der Ideen des Architekten an seine Bauarbeiter, diese führen dann den eigentlich Bauprozess durch, was in etwa der Translation gleich kommt. Die fast generelle Gültigkeit dieses Bauplans (Codes) legt nahe, dass dieser sich schon sehr früh in der Evolution des Lebens etabliert haben muss, noch vor der Abzweigung in verschiedene Organismenreiche.
Die Transkription
| Abb. 1 Die Transkription (Quelle: Google) |
Durch einen Promotor erkennt die RNA-Polymerase
den Beginn eines genetisch codierenden Abschnitts im DNA Strang, die RNA-Polymerase fährt am Matrizenstrang entlang und erzeugt so die Prä-RNA (eine unfertige Version der mRNA) diese wird dann gespleißt, was bedeutet das die nicht codierenden oder die nicht benötigten Abschnitte herausgeschnitten werden (genaueres zum Thema spleißen findet ihr in Campbell Biologie Kapitel 17). Nach erfolgreichem Spleißen erhalten wir dann die Boten-RNA (auch mRNA) diese wird dann gecapt, das bedeutet das am 5' Ende ein Guanosintriphosphat angehängt wird das verhindert das die RNA zersetzt wird. Außerdem wird am 3' Ende des Strangs eine Poly-A-Kette angehängt, eine lange Kette aus Adeninnucleoiden, dieser Poly (A)- Schwanz erleichtert den Export der mRNA aus dem Zellkern in das Cytoplasma, wo die Ribsomen (in unserem Vergleich die Bauarbeiter) die Translation (den Bau) in ein Polypeptidkette (eine lange Kette aus Aminosäuren) durchführen. Jeweils 3 Nucleotide der mRNA bilden ein Codon sozusagen einen Buchstaben. Und ein Codon ist immer einer bestimmten Aminosäure zugeordnet, Besonderheiten stellen hier noch Startcodon (AUG -> Codiert zur Aminosöure Met) und diverse Stopcodons (z.B. UGA) da. Im Unterschied zu den Eukaryoten die ich hier beschrieben habe, findet bei den Prokaryoten die Transkription direkt in eine fertige mRNA statt.
Die Translation
Sie wird durch Ribosomen durchgeführt, genauer gesagt durch eine kleine und eine große Untereinheit. Zuerst dockt die kleine Untereinheit an die Initiationsfaktoren (dem Startcodon AUG) an, danach folgen eine mit Methionin (Met) beladene transfer-RNA (t-RNA) und schließlich die große Untereinheit. Ribosomen sind eigentlich Enzyme, genannt Ribozym, sie katalysieren die Translation. Den Aufbau eines Ribosoms kann man sich in etwa so vorstellen (auch ein Ausschnitt aus der Translation):
| Abb. 2 Ribosom bei der Translation einer Polypeptidkette (Quelle: Wikipedia/Translation) |
Es gibt einen Kanal für die mRNA und 3 Bindestellen für beladene t-RNAs, die A-(Aminoacyl-), P-(Peptidyl-) und E-(Exit-)Stelle. Zuerst werden die Codons der mRNA vom Ribosom ausgelesen, welches dann die entsprechende t-RNA an der A-Stelle bindet. Im weiteren Verlauf wandert die t-RNA dann an die P-Stelle wo sich die an die t-RNA angehängt Aminosäure trennt und die t-RNA schließlich zur E-Stelle wandert wo sie wieder in das Cytoplasma entlassen wird. Durch wiederholen dieses Prozesses wächst die Polypeptidkette aus Aminosäuren immer weiter und bildet so nicht nur eine Primär- sondern meistens auch direkt eine Sekundär- und Tertiärstruktur des Proteins. Gelangt das Ribosom an ein Stopcodon so wird die Translation beendet, die Untereinheiten des Ribosoms die sich anfangs an die mRNA gebunden haben trennen sich wieder. Und fertig ist ein neues Protein.
Genaueres zum Thema Transkription und Translation findet ihr in Campbell Biologie Kap. 17. Die meisten Informationen die ich hier verwendet habe stammen ebenfalls von dort.
 |
| Abb. 3 t-RNA schematisch (Quelle: Google) |
Zum Schluss noch ein Bild zum Aufbau der t-RNA, es handelt sich dabei auch um eine lange Kette aus Nucleotiden, die durch die chemischen Bindungen untereinander, der t-RNA ihre einzigartige Form verleihen. Die verschiedenen Bereiche (auch Arme genannt) haben verschiedene Aufgaben. Der Anticodon Loop liest die komplementären Basenpaare an der mRNA und bindet an diese, während die zu codierende Aminosäure am Acceptor arm bindet.
Durch einen komplizierten Prozess der unter mitwirken von Aminoacyl-tRNA-Synthetase und ATP Verbrauch abläuft wird immer die richtige Aminosäure dem richtigen Anticodon zugeordnet. Fehler beim Lesen der ersten und zweiten Base in einem Codon können schwerwiegende Fehler, bis hin zum kompletten Ausfall des Proteins führen, Unterschiede an der dritten Position sind nicht zwangsläufig fatal.